
Chapter 7. Introduction to Cells

Chapter Outline
Type your examples here.
- 7.1 Studying Cells
- 7.2 Prokaryotic Cells
- 7.3 Eukaryotic Cells
- 7.4 The Endomembrane System
- 7.5 The Cytoskeleton
- 7.6 Connections Between Cells
Introduction
Close your eyes and picture a brick wall. What is the basic building block of that wall? A single brick, of course. Like a brick wall, your body is composed of basic building blocks, and the building blocks of your body are cells.
Your body has many kinds of cells, each specialized for a specific purpose. Just as a home is made from a variety of building materials, the human body is constructed from many cell types. For example, epithelial cells protect the surface of the body and cover the organs and body cavities within. Bone cells help to support and protect the body. Cells of the immune system fight invading bacteria. Additionally, blood and blood cells carry nutrients and oxygen throughout the body while removing carbon dioxide. Each of these cell types plays a vital role during the growth, development, and day-to-day maintenance of the body. In spite of their enormous variety, however, cells from all organisms—even ones as diverse as bacteria, onion, and human—share certain fundamental characteristics.
7.1 | Studying Cells
Learning Objectives
By the end of this section, you will be able to:
- Describe the role of cells in organisms.
- Compare and contrast light microscopy and electron microscopy.
- Summarize cell theory.
A cell is the smallest unit of a living thing. A living thing, whether made of one cell (like bacteria) or many cells (like a human), is called an organism. Thus, cells are the basic building blocks of all organisms.
Several cells of one kind that interconnect with each other and perform a shared function form tissues, several tissues combine to form an organ (your stomach, heart, or brain), and several organs make up an organ system (such as the digestive system, circulatory system, or nervous system). Several systems that function together form an organism (like a human being). Here, we will examine the structure and function of cells.
There are many types of cells, all grouped into one of two broad categories: prokaryotic and eukaryotic. For example, both animal and plant cells are classified as eukaryotic cells, whereas bacterial cells are classified as prokaryotic. Before discussing the criteria for determining whether a cell is prokaryotic or eukaryotic, let’s first examine how biologists study cells.
7.1.1 Microscopy
Most individual cells cannot be seen with the naked eye, so scientists use microscopes (micro- = “small”; -scope = “to look at”) to study them. A microscope is an instrument that magnifies an object. Most photographs of cells are taken with a microscope, and these images can also be called micrographs.
Light Microscopes
To give you a sense of cell size, a typical human red blood cell is about eight millionths of a meter, or eight micrometers (abbreviated as 8 μm) in diameter; the head of a pin of is about two thousandths of a meter (2 mm) in diameter. That means about 250 red blood cells could fit across the head of a pin.
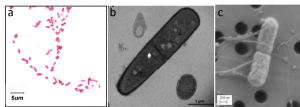
Most commonly used microscopes are classified as light microscopes (Figure 7.2a). Visible light passes and is bent through the lens system to enable the user to see the specimen. Light microscopes are advantageous for viewing living organisms, but since individual cells are generally transparent, their components are not distinguishable unless they are colored with special stains..
The optics of a light microscope’s lenses change the orientation of the image that the user sees. A specimen that is right-side up and facing right on the microscope slide will appear upside-down and facing left when viewed through a microscope, and vice versa. Similarly, if the slide is moved left while looking through the microscope, it will appear to move right, and if moved down, it will seem to move up. This occurs because microscopes use two sets of lenses to magnify the image. Because of the manner by which light travels through the lenses, this system of two lenses produces an inverted image (dissecting microscopes, work in a similar manner, but include an additional magnification system that makes the final image appear to be upright).
Light microscopes commonly used in the undergraduate college laboratory magnify up to approximately 400 times. Two parameters that are important in microscopy are magnification and resolving power. Magnification is the process of enlarging an object in appearance. Resolving power is the ability of a microscope to distinguish two adjacent structures as separate: the higher the resolution, the better the clarity and detail of the image. When oil immersion lenses are used for the study of small objects, magnification is usually increased to 1,000 times.

Electron Microscopes
In contrast to light microscopes, electron microscopes (Figure 7.2b) use a beam of electrons instead of a beam of light. Not only does this allow for higher magnification and, thus, more detail (Figure 7.3), it also provides higher resolving power. The method used to prepare the specimen for viewing with an electron microscope kills the specimen. Electrons have short wavelengths (shorter than photons) that move best in a vacuum, so living cells cannot be viewed with an electron microscope.
In a scanning electron microscope, a beam of electrons moves back and forth across a cell’s surface, creating details of cell surface characteristics. In a transmission electron microscope, the electron beam penetrates the cell and provides details of a cell’s internal structures. As you might imagine, electron microscopes are significantly more bulky and expensive than light microscopes.
![]()
Cytotechnologist
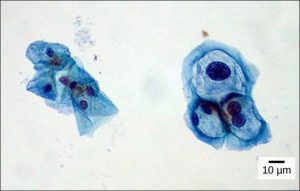
A Pap smear is a common medical test in which a small sample of cells are taken from the uterine cervix of a patient and sent to a medical lab. A professional called a cytotechnologist then stains the cells and examines them for any changes that could indicate cervical cancer or a microbial infection (Figure 7.4).
Cytotechnologists (cyto- = “cell”) study cells via microscopic examinations and other laboratory tests. They are trained to determine which cellular changes are within normal limits and which are abnormal. Their focus is not limited to cervical cells; they study cellular specimens that come from all organs. When they notice abnormalities, they consult a pathologist, who is a medical doctor who can make a clinical diagnosis. Cytotechnologists play a vital role in saving people’s lives. When abnormalities are discovered early, a patient’s treatment can begin sooner, which usually increases the chances of a successful outcome.
7.1.2 Cell Theory
The microscopes we use today are far more complex than those used in the 1600s by Antony van Leeuwenhoek, a Dutch shopkeeper who had great skill in crafting lenses. Despite the limitations of his now-ancient lenses, van Leeuwenhoek observed the movements of protista (a type of single-celled organism) and sperm, which he collectively termed “animalcules.”
In a 1665 publication called Micrographia, Robert Hooke coined the term “cell” for the box-like structures he observed when viewing cork tissue through a lens. In the 1670s, van Leeuwenhoek discovered bacteria and protozoa. Later advances in lenses, microscope construction, and staining techniques enabled other scientists to see some components inside cells. By the late 1830s, botanist Matthias Schleiden and zoologist Theodor Schwann were studying tissues and proposed the unified cell theory, which states that all living things are composed of one or more cells, the cell is the basic unit of life, and new cells arise from existing cells.
7.2 | Prokaryotic Cells
Learning Objectives
By the end of this section, you will be able to:
- Name examples of prokaryotic and eukaryotic organisms.
- Compare and contrast prokaryotic cells and eukaryotic cells.
- Describe the relative sizes of different kinds of cells.
- Explain why cells must be small.
Cells fall into one of two broad categories: prokaryotic and eukaryotic. Only the single-celled organisms of the domains Bacteria and Archaea are classified as prokaryotes. Cells of animals, plants, fungi, and protists are all eukaryotes, and are made up of eukaryotic cells.
7.2.1. Components of Prokaryotic Cells
All cells share four common components: 1) a plasma membrane, an outer covering that separates the cell’s interior from its surrounding environment; 2) cytoplasm, consisting of a jelly-like cytosol within the cell in which other cellular components are found; 3) DNA, the genetic material of the cell; and 4) ribosomes, which synthesize proteins. However, prokaryotes differ from eukaryotic cells in several ways.
A prokaryote is a simple, single-celled (unicellular) organism that lacks a nucleus, or other highly organized membrane-bound organelle (pro- = “before”; -kary- = “kernel,” which refers to the nucleus). Prokaryotic DNA is found in a central part of the cell called the nucleoid (Figure 7.5).
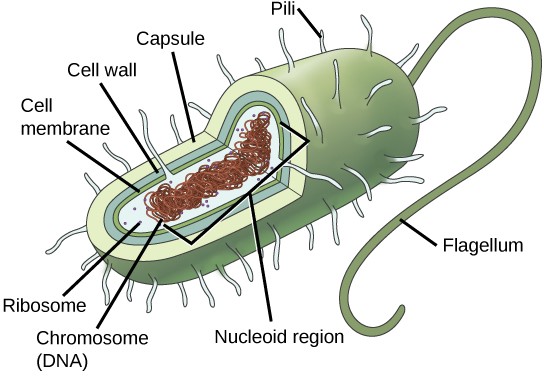
The prokaryotes, which include bacteria and archaea, are mostly single-celled organisms that, by definition, lack membrane-bound nuclei and other organelles. A bacterial chromosome is a covalently closed circle that, unlike eukaryotic chromosomes, is not organized around histone proteins. The central region of the cell in which prokaryotic DNA resides is called the nucleoid. In addition, prokaryotes often have abundant plasmids, which are shorter circular DNA molecules that may only contain one or a few genes. Plasmids can be transferred independently of the bacterial chromosome during cell division and often carry traits such as antibiotic resistance.
Most prokaryotes have a peptidoglycan cell wall and many have a polysaccharide capsule (Figure 7.5). The cell wall acts as an extra layer of protection, helps the cell maintain its shape, and prevents dehydration. The capsule enables the cell to attach to surfaces in its environment. Some prokaryotes have flagella, pili, or fimbriae. Flagella are used for locomotion. Pili are used to exchange genetic material during a type of reproduction called conjugation. Fimbriae are used by bacteria to attach to a host cell or other surface.
![]()
Microbiologist
The most effective action anyone can take to prevent the spread of contagious illnesses is to wash his or her hands. Why? Because microbes (organisms so tiny that they can only be seen with microscopes) are ubiquitous. They live on doorknobs, money, your hands, and many other surfaces. If someone sneezes into his hand and touches a doorknob, and afterwards you touch that same doorknob, the microbes from the sneezer’s mucus are now on your hands. If you touch your hands to your mouth, nose, or eyes, those microbes can enter your body and could make you sick.
However, not all microbes (also called microorganisms) cause disease; most are actually beneficial. You have microbes in your gut that make vitamin K. Other microorganisms are used to ferment beer and wine.
Microbiologists are scientists who study microbes. Microbiologists can pursue a number of careers. Not only do they work in the food industry, they are also employed in the veterinary and medical fields. They can work in the pharmaceutical sector, serving key roles in research and development by identifying new sources of antibiotics that could be used to treat bacterial infections.
Environmental microbiologists may look for new ways to use specially selected or genetically engineered microbes for the removal of pollutants from soil or groundwater, as well as hazardous elements from contaminated sites. These uses of microbes are called bioremediation technologies. Microbiologists can also work in the field of bioinformatics, providing specialized knowledge and insight for the design, development, and specificity of computer models of, for example, bacterial epidemics.
7.2.2 Cell Size
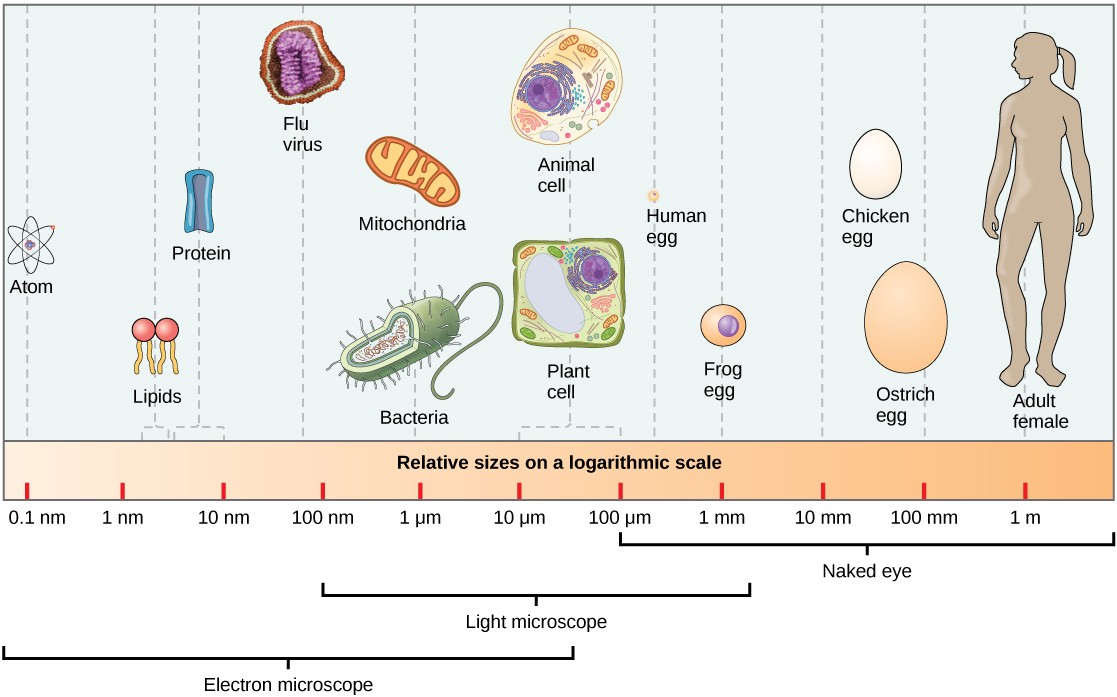
At 0.1 to 5.0 μm in diameter, prokaryotic cells are significantly smaller than most eukaryotic cells, which have diameters ranging from 10 to 100 μm (Figure 7.6). The small size of prokaryotes allows ions and organic molecules that enter them to quickly diffuse to other parts of the cell. Similarly, any wastes produced within a prokaryotic cell can quickly diffuse out. This is not the case in eukaryotic cells, which have developed different structural adaptations to enhance intracellular transport.
Small size is necessary for all cells, whether prokaryotic or eukaryotic. Not all cells are spherical in shape, but most tend to approximate a sphere. The formula for the surface area of a sphere is 4πr2, while the formula for its volume is 4πr3/3. Thus, as the radius of a cell increases, its surface area increases as the square of its radius, but its volume increases as the cube of its radius. Therefore, as a cell increases in size, its surface area-to-volume ratio decreases. This same principle would apply if the cell had the shape of a cube (Figure 7.7). If the cell grows too large, the plasma membrane will not have sufficient surface area to support the rate of diffusion required for the increased volume. In other words, as a cell grows, it becomes less efficient. One way to become more efficient is to divide; another way is to develop organelles that perform specific tasks. These adaptations lead to the development of more sophisticated cells called eukaryotic cells.
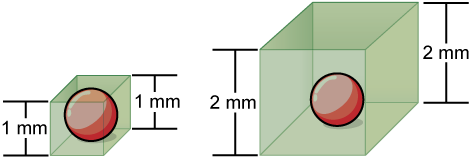
Concept Check
Prokaryotic cells are much smaller than eukaryotic cells.
- What advantages might small cell size confer on a cell?
- What advantages might large cell size have?
7.3 | Eukaryotic Cells
Learning Objectives
y the end of this section, you will be able to:
- Describe the structure of eukaryotic cells.
- Compare animal cells with plant cells.
- State the role of the plasma membrane.
- Summarize the functions of the major cell organelles.
Unlike prokaryotic cells, eukaryotic cells have a membrane-bound nucleus, numerous membrane-bound organelles that have specialized functions, and several, rod-shaped chromosomes. Because a eukaryotic cell’s nucleus is surrounded by a membrane, the word eukaryote means “true nucleus.”
Eukaryotic cells have a more complex structure than prokaryotic cells. Organelles (“little organs”) allow different functions to be compartmentalized in different areas of the cell. Before turning to organelles, let’s first examine two important components of the cell: the plasma membrane and the cytoplasm.
7.3.1 The Plasma Membrane
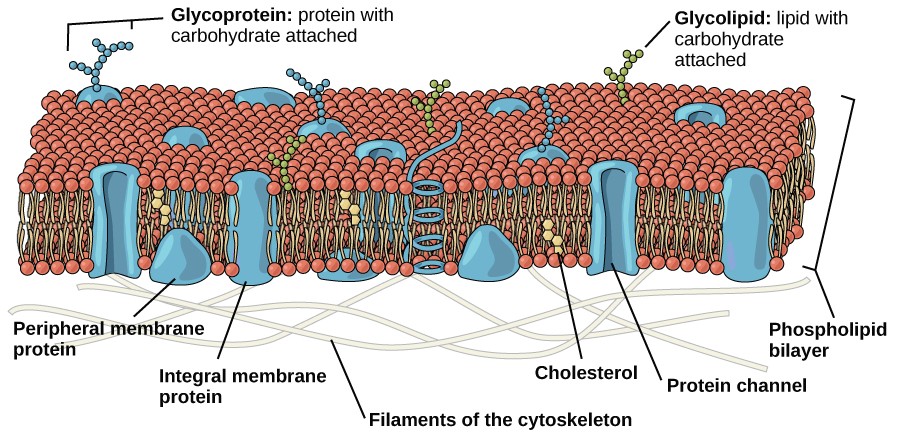
Like prokaryotes, eukaryotic cells have a plasma membrane (Figure 7.8), consisting of a phospholipid bilayer with embedded proteins, that separates the internal contents of the cell from its surrounding environment. The plasma membrane controls the passage of organic molecules, ions, water, and oxygen into and out of the cell. Wastes (such as carbon dioxide and ammonia) also leave the cell by passing through the plasma membrane.
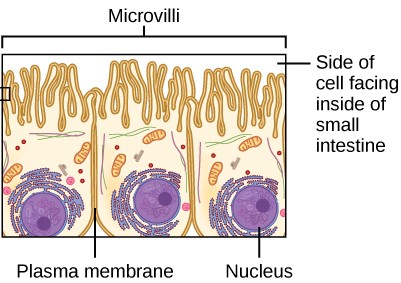
The plasma membranes of cells that specialize in absorption are folded into fingerlike projections called microvilli (Figure 7.9). Such cells are typically found lining the small intestine, the organ that absorbs nutrients from digested food. People with celiac disease have an immune response to gluten, which is a protein found in wheat, barley, and rye. The immune response damages microvilli, preventing absorption of nutrients and leading to malnutrition, cramping, and diarrhea. Patients with celiac disease must follow a gluten-free diet.
7.3.2 The Cytoplasm
The cytoplasm is the entire region of a cell between the plasma membrane and the nuclear envelope. It is made up of organelles suspended in the gel-like cytosol, the cytoskeleton, and various chemicals (Figure 7.10). Even though the cytoplasm consists of 70 to 80 percent water, it has a semi-solid consistency, which comes from the proteins within it. Glucose and other simple sugars, polysaccharides, amino acids, nucleic acids, fatty acids, and ions are also found in the cytoplasm. Many metabolic reactions take place in the cytoplasm.
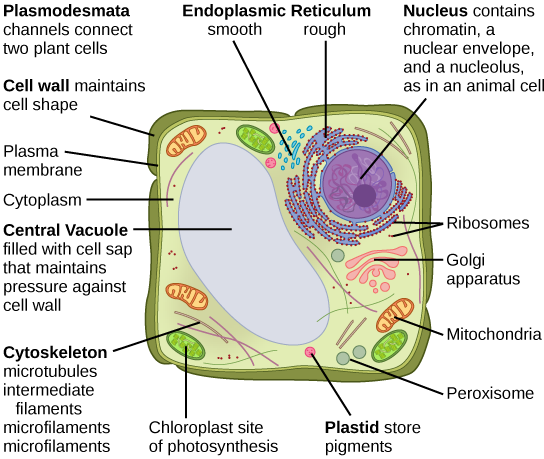
7.3.3 Organelles
The cytoplasm of eukaryotic cells is highly compartmentalized into structures called organelles. Like organs in your body, each type of organelle has a specialized function. The organelles and other cellular structures found in typical animal (Figure 7.10a) and plant (Figure 7.10b) cells are diagrammed below.

The Nucleus
Typically, the nucleus is the most prominent organelle in a cell. The nucleus (plural = nuclei) houses the cell’s DNA and directs the synthesis of ribosomes and proteins. Let’s look at it in more detail (Figure 7.11).
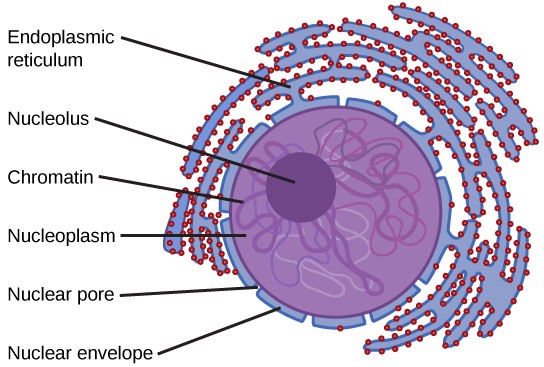
The Nuclear Envelope
The nuclear envelope is a double-membrane structure that constitutes the outermost portion of the nucleus (Figure 7.11). Both the inner and outer membranes of the nuclear envelope are phospholipid bilayers. The nuclear envelope is punctuated with pores that control the passage of ions, molecules, and RNA between the nucleoplasm and cytoplasm. The nucleoplasm is the semi-solid fluid inside the nucleus.
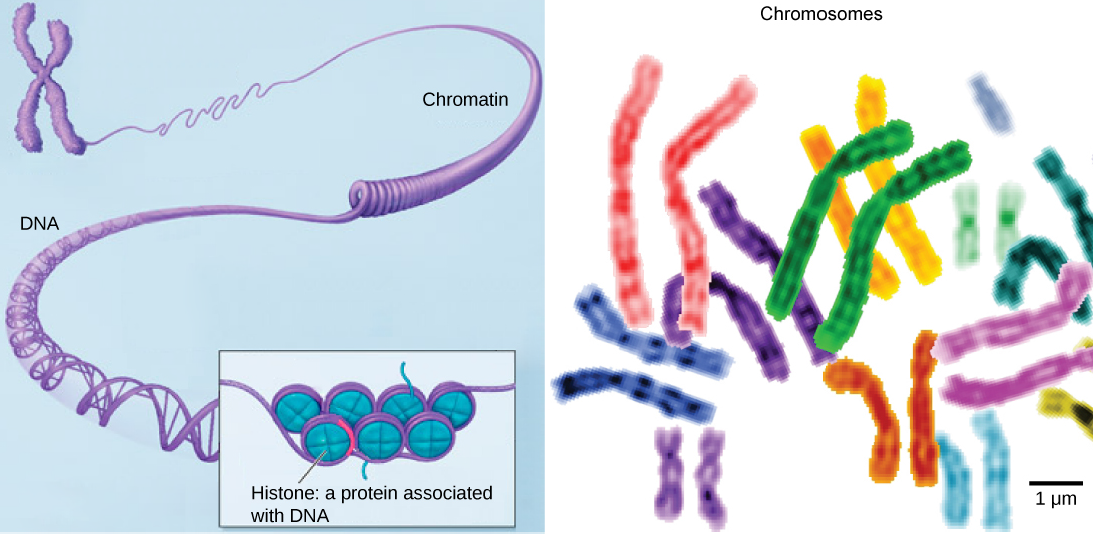
Chromatin and Chromosomes
Chromosomes are structures within the nucleus that are made up of DNA, the hereditary material. You may remember that in prokaryotes, DNA is organized into a single circular chromosome. In eukaryotes, chromosomes are linear structures. Every eukaryotic species has a specific number of chromosomes in the nuclei of its body’s cells. For example human cells have 46 chromosomes. Chromosomes are only visible and distinguishable from one another when the cell is getting ready to divide. When the cell is in the growth and maintenance phases of its life cycle, proteins are attached to chromosomes, and they resemble an unwound, jumbled bunch of threads. These unwound protein-chromosome complexes are called chromatin ( Figure 7.12).
The Nucleolus
We already know that the nucleus directs the synthesis of ribosomes, but how does it do this? Some chromosomes have sections of DNA that encode ribosomal RNA (rRNA). The darkly staining area within the nucleus called the nucleolus (plural = nucleoli) aggregates the ribosomal RNA with associated proteins to assemble ribosomal subunits that are then transported out through the pores in the nuclear envelope to the cytoplasm.
Ribosomes
Ribosomes are the cellular structures responsible for protein synthesis. They are large complexes of protein and rRNA, consisting of a large and a small subunit (Figure 7.13). When viewed through an electron microscope, ribosomes appear either as clusters or as single, tiny dots that float freely in the cytoplasm. They may be attached to the inside of the plasma membrane or the outside of the endoplasmic reticulum or nuclear envelope (Figure 7.11). Ribosomes bind to mRNA and translate the code provided by the sequence of the nitrogenous bases in the mRNA into a specific order of amino acids in a protein.
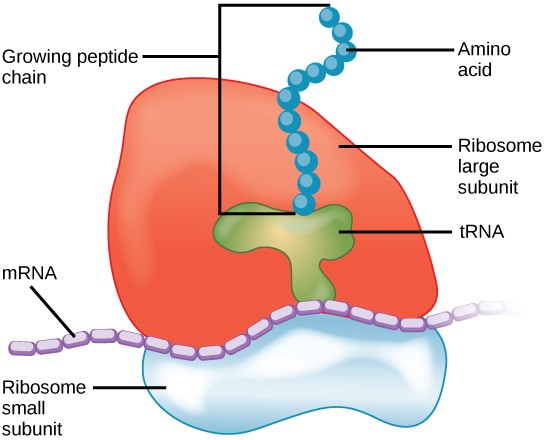
Because proteins synthesis is an essential function of all cells, ribosomes are found in practically every cell. Ribosomes are particularly abundant in cells that synthesize large amounts of protein. For example, the pancreas is responsible for creating several digestive enzymes and the cells that produce these enzymes contain many ribosomes.
Mitochondria
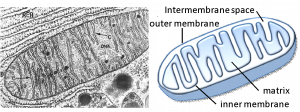
Mitochondria (singular = mitochondrion) are often called the “powerhouses” or “energy factories” of a cell because they are responsible for making adenosine triphosphate (ATP), the cell’s main energy-carrying molecule. ATP is made using the chemical energy found in glucose and other nutrients by the process of cellular respiration. In mitochondria, this process uses oxygen and produces carbon dioxide as a waste product. In fact, the carbon dioxide that you exhale with every breath comes from the breakdown of nutrients you eat inside mitochondria.
Mitochondria are oval-shaped, double membrane organelles (Figure 7.14) that have their own ribosomes and DNA. Each membrane is a phospholipid bilayer embedded with proteins. The inner membrane has folds called cristae that increase surface area. The space between the two membranes is called the intermembrane space and the space inside the inner membrane is called the matrix. ATP synthesis takes place on the inner membrane.
Peroxisomes
Peroxisomes are small, spherical organelles enclosed by single membranes. They carry out oxidation reactions that break down fatty acids and amino acids. They also detoxify many poisons that may enter the body. (Many of these oxidation reactions release hydrogen peroxide, H2O2, which would be damaging to cells; however, when these reactions are confined to peroxisomes, enzymes safely break down the H2O2 into oxygen and water.) For example, alcohol is detoxified by peroxisomes in liver cells. Glyoxysomes, which are specialized peroxisomes in plants, are responsible for converting stored fats into sugars.
Vesicles and Vacuoles
Vesicles and vacuoles are membrane-bound sacs that function in storage and transport. Other than the fact that vacuoles are somewhat larger than vesicles, there is a very subtle distinction between them: The membranes of vesicles can fuse with either the plasma membrane or other membrane systems within the cell. Additionally, some agents such as enzymes within plant vacuoles break down macromolecules. The membrane of a vacuole does not fuse with the membranes of other cellular components. Some types of cells have contractile vacuoles, which contract to expel water from the cell.
7.,3.4 Animal Cells versus Plant Cells
At this point, you know that each eukaryotic cell has a plasma membrane, cytoplasm, a nucleus, ribosomes, mitochondria, peroxisomes, and in some, vacuoles, but there are some striking differences between animal and plant cells. While both animal and plant cells have microtubule organizing centers (MTOCs), animal cells also have centrioles associated with the MTOC: a complex called the centrosome. Animal cells each have a centrosome and lysosomes, whereas plant cells do not. Plant cells have a cell wall, chloroplasts and other specialized plastids, and a large central vacuole, whereas animal cells do not.
The Centrosome
The centrosome is a microtubule-organizing center from which all microtubules in the cell originate. It is found near the nucleus of animal cells. Each centrosome contains a pair of centrioles, which lie perpendicular to each other (Figure 7.15). Each centriole is a cylinder of nine triplets of microtubules.
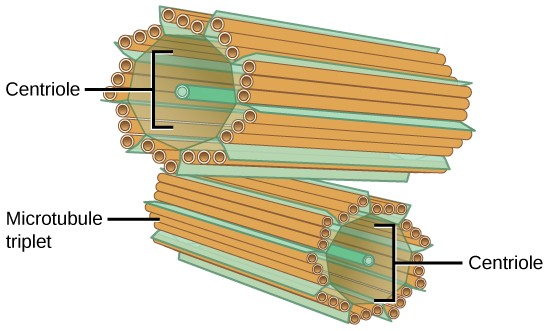
The centrosome replicates itself before a cell divides, and the centrioles appear to have some role in pulling the duplicated chromosomes to opposite ends of the dividing cell. However, the exact function of the centrioles in cell division isn’t clear, because cells that have had the centrosome removed can still divide, and plant cells, which lack centrosomes, are capable of cell division.
Lysosomes
Animal cells have another set of organelles not found in plant cells: lysosomes. Lysosomes are the cell’s “garbage disposal.” Enzymes inside lysosomes aid the breakdown of proteins, polysaccharides, lipids, nucleic acids, and even worn- out organelles. These enzymes are active at a much lower pH than that of the cytoplasm. Therefore, the pH within lysosomes is more acidic than the pH of the cytoplasm. Proton pumps in the lysosome membrane pump protons in to maintain the low pH inside the lysosome. In plant cells, digestive processes take place in vacuoles.
The Cell Wall
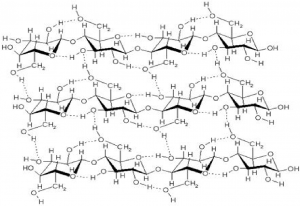
If you examine Figure 7.10b, the diagram of a plant cell, you will see a structure external to the plasma membrane called the cell wall. The cell wall is a rigid covering that protects the cell, provides structural support, and gives shape to the cell. While the chief component of prokaryotic cell walls is peptidoglycan, the major organic molecule in the plant cell wall is cellulose (Figure 7.16), a polysaccharide made up of glucose units. When you bite into a raw vegetable, like celery, it crunches because you are tearing the rigid cell walls of the celery cells with your teeth. Fungus and some protist cells also have cell walls, which are made of other structural molecules.
Chloroplasts
Like mitochondria, chloroplasts have their own DNA and ribosomes, but chloroplasts have an entirely different function. Chloroplasts are plant cell organelles that carry out photosynthesis. Photosynthesis is the series of reactions that use carbon dioxide, water, and light energy to make glucose and oxygen. This is a major difference between plants and animals; plants (autotrophs) are able to make their own food, like sugars, while animals (heterotrophs) must ingest their food.
Like mitochondria, chloroplasts have outer and inner membranes, but within the space enclosed by a chloroplast’s inner membrane is a set of interconnected and stacked fluid-filled membrane sacs called thylakoids (Figure 7.17). Each stack of thylakoids is called a granum (plural = grana). The fluid enclosed by the inner membrane that surrounds the grana is called the stroma. The fluid inside the thylakoids is called the thylakoid space. Light energy is harvested in the thylakoid membranes and sugar is made in the stroma.
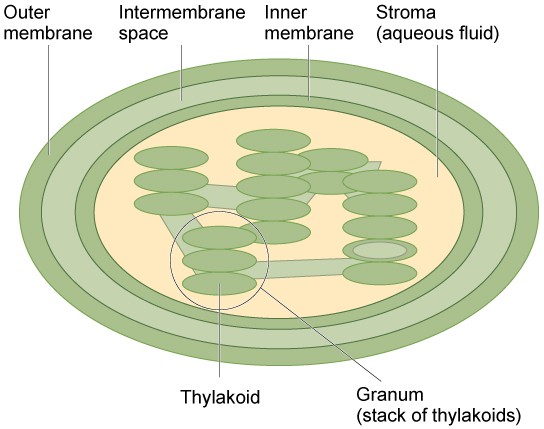
Chloroplasts contain a green pigment called chlorophyll, which captures the light energy that drives the reactions of photosynthesis. Like plant cells, photosynthetic protists also have chloroplasts. Some bacteria perform photosynthesis, but their photosynthetic pigments are not relegated to an organelle.
The Central Vacuole
Plant cells each have a large central vacuole that occupies most of the area of the cell (Figure 7.8b). The central vacuole plays a key role in regulating the cell’s concentration of water in changing environmental conditions. If you forget to water a plant for a few days, it wilts because when environmental water levels are low, water moves out of the central vacuoles and cytoplasm. As the central vacuole shrinks, it leaves the cell wall unsupported, resulting in wilting. The central vacuole also supports the expansion of the cell. When the central vacuole holds more water, the cell gets larger without having to invest a lot of energy in synthesizing new cytoplasm.

Endosymbiosis
We have mentioned that both mitochondria and chloroplasts contain DNA and ribosomes. Have you wondered why? Strong evidence points to endosymbiosis as the explanation.
Symbiosis is a relationship in which organisms from two separate species depend on each other for their survival. Endosymbiosis (endo- = “within”) is a mutually beneficial relationship in which one organism lives inside the other. Endosymbiotic relationships abound in nature. We have already mentioned that microbes that produce vitamin K live inside the human gut. This relationship is beneficial for us because we are unable to synthesize vitamin K. It is also beneficial for the microbes because they are protected from other organisms and from drying out, and they receive abundant food from the environment of the large intestine.
Scientists have long noticed that bacteria, mitochondria, and chloroplasts are similar in size. We also know that bacteria have DNA and ribosomes, just as mitochondria and chloroplasts do. Scientists believe that host cells and bacteria formed an endosymbiotic relationship when the host cells ingested both aerobic and autotrophic bacteria (cyanobacteria) but did not destroy them. Through many millions of years of evolution, these ingested bacteria became more specialized in their functions, with the aerobic bacteria becoming mitochondria and the autotrophic bacteria becoming chloroplasts.
7.4 | The Endomembrane System
Learning Objectives
By the end of this section, you will be able to:
- List the components of the endomembrane system.
- Recognize the relationship between the endomembrane system and its functions.
The endomembrane system (endo = “within”) is a group of membranes and organelles (Figure 7.18) in eukaryotic cells that works together to modify, package, and transport lipids and proteins. It includes the nuclear envelope, lysosomes, and vesicles, which we’ve already mentioned, as well as the endoplasmic reticulum and Golgi apparatus. Although not technically within the cell, the plasma membrane is sometimes included in the endomembrane system because it interacts with the other endomembranous organelles.
7.4.1 The Endoplasmic Reticulum
The endoplasmic reticulum (ER) is a series of interconnected membranous sacs and tubules that collectively modifies proteins and synthesizes lipids. However, these two functions are performed in separate areas of the ER: the rough ER and the smooth ER, respectively.
The hollow portion of the ER tubules is called the lumen or cisternal space. The membrane of the ER, which is a phospholipid bilayer embedded with proteins, is continuous with the nuclear envelope.
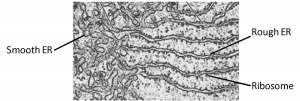
Rough ER
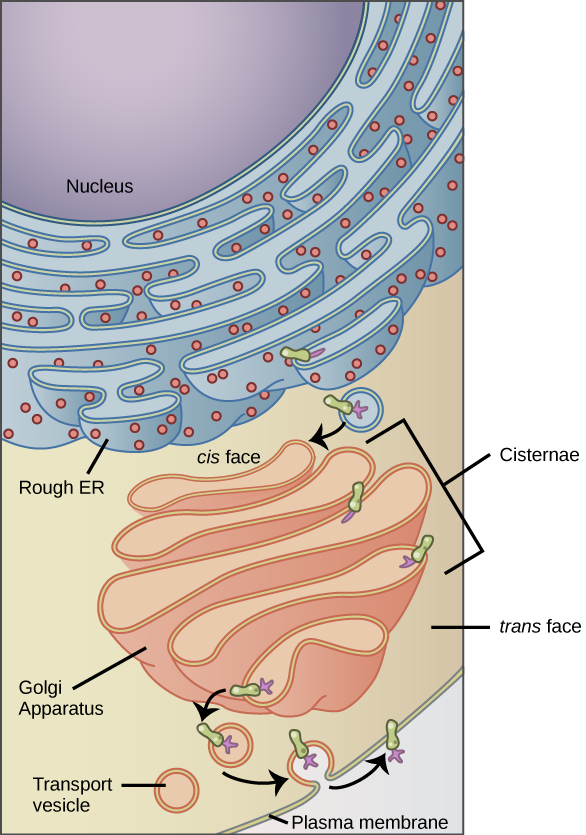
The rough endoplasmic reticulum (RER) is so named because ribosomes attached to its cytoplasmic surface give it a studded appearance when viewed through an electron microscope (Figure 7.19). These bound ribosomes are in the process of translating proteins directly into the lumen of the RER.
Three types of proteins are made on bound ribosomes: proteins that will end up secreted from the cell: proteins that will end up inserted into the plasma membrane of the cell, and proteins that will end up inside organelles in the cell. What these three classes of proteins have in common is that they will not end up in the cytosol of the cell. From the moment that they start to be synthesized, these proteins are sequestered inside membrane-bound compartments and will never touch the cytosol.
Once the newly synthesized proteins are deposited into the lumen of the RER, they undergo structural modifications, such as folding or addition of side chains. If the modified proteins are not destined to stay in the RER, they will travel by small, membrane-bound transport vesicles that bud from the RER’s membrane to the Golgi apparatus (Figure 7.18).
Since the RER is engaged in modifying proteins that will be secreted from the cell, the RER is abundant in cells that secrete proteins, such as liver cells, for example.
Smooth ER
The smooth endoplasmic reticulum (SER) is continuous with the RER but has few or no ribosomes on its surface (Figure 7.19). Functions of the SER include synthesis of carbohydrates, lipids, phospholipids, and steroid hormones and detoxification of medications and poisons. The SER takes on different functions depending on the needs of the cell. For example, in muscle cells, a specialized SER called the sarcoplasmic reticulum stores calcium ions that are needed to trigger the coordinated contractions of the muscle cells. Cells that make a lot of lipids have a large amount of SER. For example, Leydig cells in mammalian testes produce steroid hormones such as testosterone and therefore have abundant SER.
7.4.2 The Golgi Apparatus
Vesicles containing proteins and lipids bud from the ER and transport their contents to the Golgi apparatus (GA). In the GA, proteins and lipids are sorted, packaged, and tagged so that they wind up in the right place. For this reason, the GA is sometimes called the post office of the cell. The GA is a series of flattened membranes, each forming a separate compartment The receiving side of the Golgi apparatus is called the cis face. The opposite side is called the trans face (Figure 7.20).
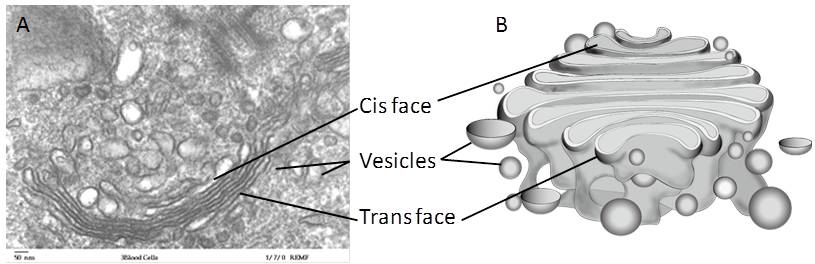
Transport vesicles from the ER fuse with the cis face and empty their contents into the lumen of the GA. The proteins and lipids travel from compartment to compartment by vesicle. As they travel through the GA, they undergo further modifications. The most frequent modification is the addition of short chains of sugar molecules. They may also be tagged with phosphate groups or other small molecules so that they can be routed to their proper destinations.
Finally, the modified and tagged proteins are packaged into vesicles that bud from the trans face of the Golgi. Some of these vesicles deposit their contents into other organelles of the cell. Other vesicles fuse with the plasma membrane and their contents end up outside the cell or inserted into the plasma membrane.
Cells that engage in a great deal of secretory activity, such as cells of the salivary glands that secrete digestive enzymes, have an abundance of GA. In plant cells, the Golgi apparatus has the additional role of synthesizing polysaccharides, some of which are incorporated into the cell wall and some of which are used in other parts of the cell.
![]()
Cardiologist
Heart disease is the leading cause of death in the United States. This is primarily due to sedentary lifestyles and poor diets. Heart failure is just one of many disabling heart conditions. Heart failure occurs when the heart cannot pump with sufficient force to transport oxygenated blood to all the vital organs.
Heart failure occurs when the endoplasmic reticula of cardiac muscle cells do not function properly. As a result, an insufficient number of calcium ions are available to trigger a sufficient contractile force.
Cardiologists (cardi- = “heart”) are doctors who specialize in treating heart diseases. Cardiologists can diagnose heart failure via physical examination, results from an electrocardiogram (a test that measures the electrical activity of the heart), a chest X-ray to see whether the heart is enlarged, and other tests. If heart failure is diagnosed, the cardiologist will typically prescribe appropriate medications and recommend a reduction in table salt intake and a supervised exercise program.
7.4.3 Lysosomes
Lysosomes are also part of the endomembrane system. Lysosomes are small, spherical compartments that function as the digestive and organelle-recycling facility of animal cells, They contain hydrolytic enzymes that digest non-functioning organelles, macromolecules, and pathogens (disease-causing organisms). The enzymes in lysosomes are called acid hydrolases because they catalyze hydrolysis reaction and their optimal pH is around 4.5. Lysosomes contain proton pumps in their membranes that pump hydrogen ions into the lumen, thereby lowering the pH. This elegant mechanism allows the enzymes to be safely made and transported through the endomembrane system in an inactive state. They become active only when they encounter the acidic environment inside the lysosome.
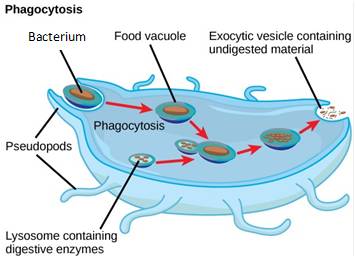
Lysosomes also use their hydrolytic enzymes to destroy pathogens that enter the cell. A good example of this occurs in a type of white blood cells called macrophages, which are part of your body’s immune system. In a process known as phagocytosis, macrophages engulf pathogens into a vesicle. The vesicle containing the pathogen fuses with a lysosome, and the lysosome’s enzymes destroy the pathogen (Figure 7.21).
7.5 | The Cytoskeleton
Learning Objectives
By the end of this section, you will be able to:
- Describe the cytoskeleton.
- Compare the roles of microfilaments, intermediate filaments, and microtubules.
- Compare and contrast cilia and flagella.
- Summarize the differences among the components of prokaryotic cells, animal cells, and plant cells.
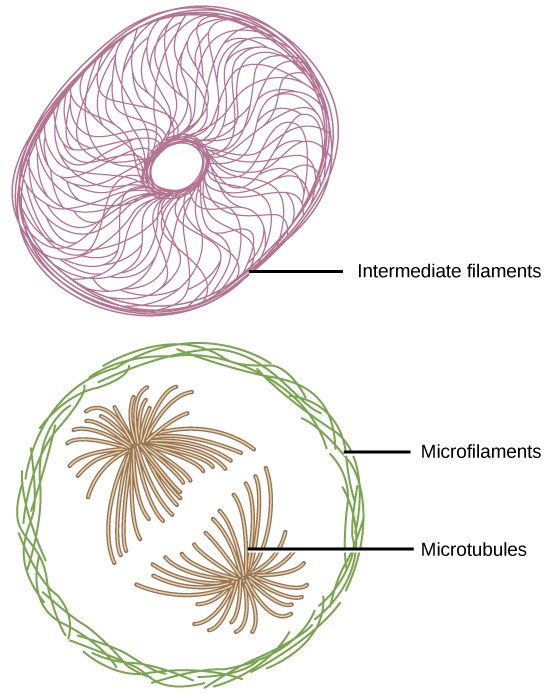
If you were to remove all the organelles from a cell, would the plasma membrane and the cytoplasm be the only components left? No. Within the cytoplasm, there would still be ions and organic molecules, plus a network of protein fibers that help maintain the shape of the cell, secure some organelles in specific positions, allow cytoplasm and vesicles to move within the cell, and enable cells within multicellular organisms to move. Collectively, this network of protein fibers is known as the cytoskeleton. There are three types of fibers within the cytoskeleton: microfilaments, intermediate filaments, and microtubules (Figure 7.22). Here, we will examine each.
7.5.1 Microfilaments
Of the three types of protein fibers in the cytoskeleton, microfilaments are the thinnest. They function in cellular movement and are made of two intertwined strands of a globular protein called actin (Figure 7.23). For this reason, microfilaments are also known as actin filaments.
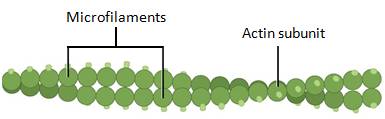
Microfilaments provide some rigidity and shape to cells. They can depolymerize (disassemble) and reform quickly, thus enabling a cell to change its shape and move. Microfilaments are also involved in cellular movement, such as cell division in animal cells and cytoplasmic streaming, which is the circular movement of the cell cytoplasm, in plant cells. Actin also helps muscle cells contract.
7.5.2 Intermediate Filaments
Intermediate filaments are made of several strands of fibrous proteins that are wound together (Figure 7.24). These elements of the cytoskeleton get their name from the fact that their diameter is between those of microfilaments and microtubules. The function of intermediate filaments is purely structural. They bear tension, thus maintaining the shape of the cell, and create a supportive scaffolding to anchor the nucleus and other organelles in place.
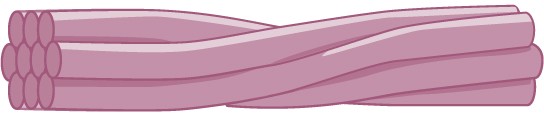
Intermediate filaments are the most diverse group of cytoskeletal elements. Several types of fibrous proteins are found in the intermediate filaments, including keratin, the fibrous protein that strengthens your hair, nails, and skin.
7.5.3 Microtubules
As their name implies, microtubules are small hollow tubes. The walls of the microtubule are made of dimers of α-tubulin and β-tubulin, two globular proteins (Figure 7.25). Microtubules are the widest components of the cytoskeleton. They help the cell resist compression, provide a track along which vesicles move through the cell, and pull replicated chromosomes to opposite ends of a dividing cell. Like microfilaments, microtubules can dissolve and reform quickly.
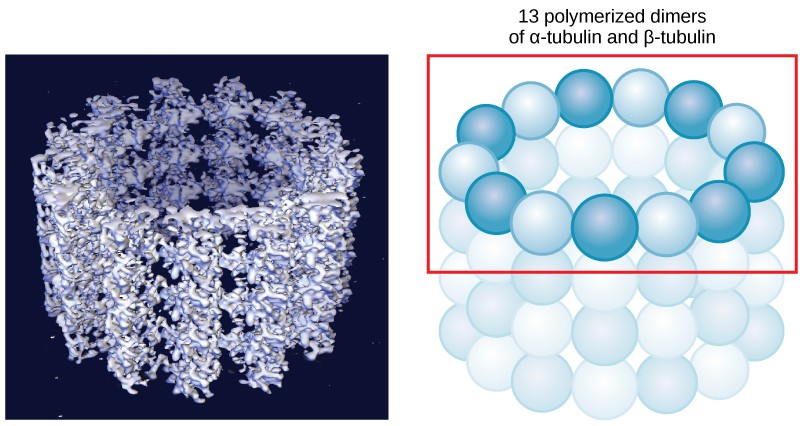
Microtubules are also the structural elements of flagella, cilia, and centrioles. In fact, in animal cells, the centrioles are the microtubule-organizing center. In eukaryotic cells, flagella and cilia are quite different structurally from their counterparts in prokaryotes, as discussed below.
Flagella and Cilia
Flagella (singular = flagellum) are long, hair-like structures that extend from the plasma membrane and are used to move an entire cell. When present, a cell has just one flagellum or a few flagella. Cilia (singular = cilium) are short, hair-like structures that are used to move entire cells or to move substances along the outer surface of the cell. For example, the cells lining the ovarian tubes have cilia that move the ovum toward the uterus, and the cells lining the respiratory tract have cilia that move mucus toward your nostrils. When cilia are present, they extend along the entire surface of the plasma membrane.
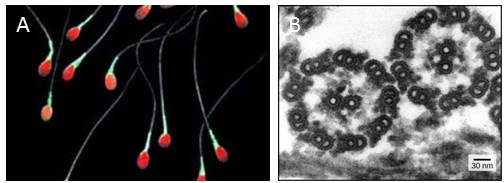
Despite their differences in length and number, flagella and cilia share a common structural arrangement of microtubules called a “9 + 2 array.” A single flagellum or cilium is made of a ring of nine microtubule pairs surrounding a single microtubule pair in the center (Figure 7.26).
Table 7.1 Components of prokaryotic and eukaryotic cells.
|
Component |
Function |
In Pro-karyotes? |
In Animal Cells? |
In Plant Cells? |
|
Plasma membrane |
Separates cell from external environment; controls passage of organic molecules, ions, water, oxygen, and wastes in and out of cell |
Yes |
Yes |
Yes |
|
Cytoplasm |
Provides turgor pressure to plant cells as fluid inside the central vacuole; site of many metabolic reactions; medium in which organelles are found |
Yes |
Yes |
Yes |
|
Nucleus |
Cell organelle that houses DNA and directs synthesis of ribosomes and proteins |
No |
Yes |
Yes |
|
Nucleolus |
Darkened area within the nucleus where ribosomal subunits are synthesized. |
No |
Yes |
Yes |
|
Ribosomes |
Protein Synthesis |
Yes |
Yes |
Yes |
|
Mitochondria |
ATP production/cellular respiration |
No |
Yes |
Yes |
|
Peroxisomes |
Oxidizes and thus breaks down fatty acids and amino acids, and detoxifies poisons |
No |
Yes |
Yes |
|
Vesicles and Vacuoles |
Storage and transport; digestive function in plant cells |
No |
Yes |
Yes |
|
Centrosome |
Unspecified role in cell division in animal cells; source of microtubules in animal cells |
No |
Yes |
No |
|
Lysosome |
Digestion of macromolecules; recycling of worn-out organelles |
No |
Yes |
No |
|
Cell Wall |
Protection, structural support and maintenance of cell shape |
Yes |
No |
Yes |
|
Chloroplasts |
Photosynthesis |
No |
No |
Yes |
|
Endoplasmic Reticulum |
Modifies proteins and synthesizes lipids |
No |
Yes |
Yes |
|
Golgi Apparatus |
Modifies, sorts, tags, packages, and distributes lipids and proteins |
No |
Yes |
Yes |
|
Cytoskeleton |
Maintains cell’s shape, secures organelles in specific positions, allows cytoplasm and vesicles to move within cell, and enables unicellular organisms to move independently |
Yes |
Yes |
Yes |
|
Flagella |
Cellular locomotion |
Some |
Some |
Rarely – in sperm |
|
Cilia |
Cellular locomotion, movement of particles along extracellular surface of plasma membrane, and filtration |
Some |
Some |
No |
7.6 | Connections Between Cells
Learning Objectives
By the end of this section, you will be able to:
- Describe the extracellular matrix.
- List examples of the ways that plant cells and animal cells communicate with adjacent cells.
- Summarize the roles of tight junctions, desmosomes, gap junctions, and plasmodesmata.
You already know that a group of similar cells working together is called a tissue. As you might expect, if cells are to work together, they must communicate with each other, just as you need to communicate with others if you work on a group project. Let’s take a look at how cells communicate with each other.
7.6.1 Extracellular Matrix of Animal Cells
Most animal cells release materials into the extracellular space. The primary components of these materials are proteins, and the most abundant protein is collagen. Collagen fibers are interwoven with carbohydrate-containing protein molecules called proteoglycans. Collectively, these materials are called the extracellular matrix (Figure 7.27). Not only does the extracellular matrix hold the cells together to form a tissue, but it also allows the cells within the tissue to communicate with each other. How can this happen?
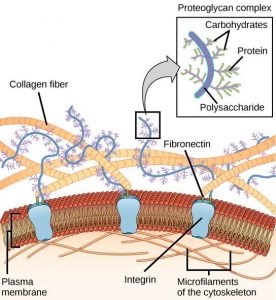
Cells have protein receptors on the extracellular surfaces of their plasma membranes. When a molecule within the matrix binds to the receptor, it changes the molecular structure of the receptor. The receptor, in turn, changes the conformation of the microfilaments positioned just inside the plasma membrane. These conformational changes induce chemical signals inside the cell that reach the nucleus and turn “on” or “off” the transcription of specific sections of DNA, which affects the production of associated proteins, thus changing the activities within the cell.
Blood clotting provides an example of the role of the extracellular matrix in cell communication. When the cells lining a blood vessel are damaged, they display a protein receptor called tissue factor. When tissue factor binds with another factor in the extracellular matrix, it causes platelets to adhere to the wall of the damaged blood vessel, stimulates the adjacent smooth muscle cells in the blood vessel to contract (thus constricting the blood vessel), and initiates a series of steps that stimulate the platelets to produce clotting factors.
7.6.2 Intercellular Junctions
Adjacent cells can also communicate with each directly through intercellular junctions. Plasmodesmata are junctions between plant cells; tight junctions, gap junctions, and desmosomes occur between animal cells.
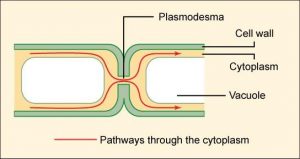
Plasmodesmata
In general, plasma membranes of neighboring plant cells cannot touch one another because they are separated by the cell wall that surrounds each cell. Plasmodesmata (singular = plasmodesma) are numerous channels that pass between cell walls of adjacent plant cells, connect their cytoplasm, and enable materials to be transported from cell to cell (Figure 7.28). This allows plants to transfer water and other soil nutrients from its roots, through its stems, and to its leaves.
Tight Junctions
A tight junction is a watertight seal between two adjacent animal cells (Figure 7.29A). The cells are held tightly against each other by proteins, preventing materials from leaking between the cells. Tight junctions are typically found in tissues that line internal organs and cavities. For example, the tight junctions of the epithelial cells lining your urinary bladder prevent urine from leaking out into the extracellular space. Tight junctions in the gut normally prevent food from leaking between cells into your body. However, these junctions can be regulated to allow some substances to pass through.
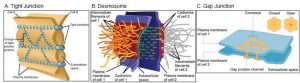
Desmosomes
Also found only in animal cells are desmosomes, which act like spot welds between adjacent epithelial cells (Figure 7.29B). Proteins in the plasma membrane connect to intermediate filaments to create desmosomes. Effectively, the cytoskeletons of the two cells are linked together so they cannot easily be pulled apart. Skin is an example of a tissue with numerous desmosomes. This allows you to pull on your skin without it ripping apart.
Gap Junctions
Gap junctions in animal cells are similar to plant cell plasmodesmata. They are channels between adjacent cells that allow for the transport of ions, nutrients, and other substances that enable cells to communicate (Figure 7.29C). Structurally, however, gap junctions and plasmodesmata differ.
Gap junctions develop when a set of six proteins (called connexins) in the plasma membrane arrange themselves in a ring, called a connexon. When the pores of connexons in adjacent cells align, they form a channel between the two cells. Gap junctions are particularly important in cardiac muscle. The electrical signal for the muscle to contract is passed efficiently through gap junctions, allowing the heart muscle cells to contract in unison.
